







Research teams
- Seed Development
- Epigenetics, Chromatin and Development
- Evolution and Development of the Flower
- Mechanotransduction in Development
- Floral Morphogenesis
- Cell Signalling and Endomembranes
- Hormonal Signalling and Development
- MOrphogenesis Simulation and Analysis In siliCo (MOSAIC)
- [Bioinformatics Hub]
 Charlotte KIRCHHELLE
Charlotte KIRCHHELLE
CR INRAE
contact
Publications
 Olivier HAMANT
Olivier HAMANT
DR INRAE
contact
Publications
 Christophe TREHIN
Christophe TREHIN
MC UCBL
contact
 Marjolaine MARTIN
Marjolaine MARTIN
TR ENS
contact
 Claire Lionnet
Claire Lionnet
IE CNRS
contact
 Annalisa BELLANDI
Annalisa BELLANDI
EMBO Long-Term Fellow
contact
 Antoine CHEVALLIER
Antoine CHEVALLIER
PhD student
contact
 Denise ARICO
Denise ARICO
Postdoc INRAE
contact
 Isaty MELOGNO
Isaty MELOGNO
PhD student
contact
 Johanna DICKMANN
Johanna DICKMANN
Walter Benjamin fellow (DFG)
contact
 Liam ELLIOTT
Liam ELLIOTT
Postdoc INRAE
contact
 Mariana ROMEIRO MOTTA
Mariana ROMEIRO MOTTA
Postdoc INRAE
contact
 Marianne LANG
Marianne LANG
PhD student
contact
 Nathan GERMAN
Nathan GERMAN
PhD Student
contact
 Sana DIEUDONNE
Sana DIEUDONNE
IR INRAE
contact
 Zoe NEMEC VENZA
Zoe NEMEC VENZA
Postdoc INRAE
contact
2025
Journal articles
- titre
- Why participatory plant research now?
- auteur
- Sofía Correa, Marie-Thérèse Charreyre, Olivier Hamant, Mathieu Thomas
- article
- Quantitative Plant Biology, 2025, 6, pp.e4. ⟨10.1017/qpb.2024.20⟩
- Accès au texte intégral et bibtex
-


Preprints, Working Papers, ...
- titre
- A Rab-Kinesin12-Fused kinase module couples vesicle delivery and phragmoplast remodelling during cytokinesis in Arabidopsis
- auteur
- Liam Elliott, Monika Kalde, Moé Yamada, Michal Hála, Andrei Smertenko, Frédérique Rozier, James Evry, Niloufer Irani, Yvon Jaillais, Patrick Hussey, Viktor Žárský, Ian Moore, Charlotte Kirchhelle
- article
- 2025
- Accès au bibtex
-
2024
Journal articles
- titre
- Northern Normal: Laboratory Networks, Microbial Culture Collections, and Taxonomies of Power (1939–2000)
- auteur
- Claas Kirchhelle, Charlotte Kirchhelle
- article
- Engaging Science, Technology, and Society, 2024, 10 (1–2), ⟨10.17351/ests2023.1445⟩
- Accès au texte intégral et bibtex
-
- titre
- A 3-component module maintains sepal flatness in Arabidopsis
- auteur
- Shouling Xu, Xi He, Duy-Chi Trinh, Xinyu Zhang, Xiaojiang Wu, Dengying Qiu, Ming Zhou, Dan Xiang, Adrienne H.K. Roeder, Olivier Hamant, Lilan Hong
- article
- Current Biology, 2024, 34 (17), pp.4007-4020.e4. ⟨10.1016/j.cub.2024.07.066⟩
- Accès au texte intégral et bibtex
-
- titre
- Arabidopsis floral buds are locked through stress-induced sepal tip curving
- auteur
- Duy-Chi Trinh, Isaty Melogno, Marjolaine Martin, Christophe Trehin, Richard S Smith, Olivier Hamant
- article
- Nature Plants, 2024, 10 (8), pp.1258-1266. ⟨10.1038/s41477-024-01760-6⟩
- Accès au texte intégral et bibtex
-
- titre
- Sepal shape variability is robust to cell size heterogeneity in Arabidopsis
- auteur
- Duy-Chi Trinh, Claire Lionnet, Christophe Trehin, Olivier Hamant
- article
- Biology Letters, 2024, 20 (5), pp.20240099. ⟨10.1098/rsbl.2024.0099⟩
- Accès au texte intégral et bibtex
-
- titre
- Contributions of lignification, tissue arrangement patterns, and cross-sectional area to whole-stem mechanical properties in Arabidopsis thaliana
- auteur
- Mariko Asaoka, Eric Badel, Ali Ferjani, Kazuhiko Nishitani, Olivier Hamant
- article
- Journal of Plant Research, 2024, 137 (5), pp.773-783. ⟨10.1007/s10265-024-01543-2⟩
- Accès au bibtex
-
- titre
- Debunking the idea of biological optimisation: quantitative biology to the rescue
- auteur
- Olivier Hamant
- article
- Quantitative Plant Biology, 2024, 5, pp.e3. ⟨10.1017/qpb.2024.3⟩
- Accès au texte intégral et bibtex
-
- titre
- Unraveling gluten protein compositions of einkorn, emmer, and spelt grown in the Lyon region in France
- auteur
- Sofía Correa, Justine Lepagneul, Mathieu Thomas, Laurence Mayaud, Olivier Hamant, Marie-Françoise Samson, Marie‐thérèse Charreyre
- article
- Cereal Chemistry, 2024, 101 (4), pp.798-816. ⟨10.1002/cche.10781⟩
- Accès au bibtex
-
- titre
- Guidelines for naming and studying plasma membrane domains in plants
- auteur
- Yvon Jaillais, Emmanuelle Bayer, Dominique Bergmann, Miguel Botella, Yohann Boutté, Tolga Bozkurt, Marie-Cecile Caillaud, Véronique Germain, Guido Grossmann, Ingo Heilmann, Piers Hemsley, Charlotte Kirchhelle, Alexandre Martinière, Yansong Miao, Sebastien Mongrand, Sabine Müller, Lise Noack, Yoshihisa Oda, Thomas Ott, Xue Pan, Roman Pleskot, Martin Potocky, Stéphanie Robert, Clara Sanchez Rodriguez, Françoise Simon-Plas, Eugenia Russinova, Daniel van Damme, Jaimie van Norman, Dolf Weijers, Shaul Yalovsky, Zhenbiao Yang, Enric Zelazny, Julien Gronnier
- article
- Nature Plants, 2024, 10 (8), pp.1172-1183. ⟨10.1038/s41477-024-01742-8⟩
- Accès au texte intégral et bibtex
-
- titre
- A transient radial cortical microtubule array primes cell division in Arabidopsis
- auteur
- Isaty Melogno, Shogo Takatani, Paula Llanos, Coralie Goncalves, Chie Kodera, Marjolaine Martin, Claire Lionnet, Magalie Uyttewaal, Martine Pastuglia, Christophe Trehin, David Bouchez, Jacques Dumais, Olivier Hamant
- article
- Proceedings of the National Academy of Sciences of the United States of America, 2024, 121 (29), pp.e2320470121. ⟨10.1073/pnas.2320470121⟩
- Accès au texte intégral et bibtex
-
- titre
- Is incoherence required for sustainability?
- auteur
- Olivier Hamant
- article
- Anthropocene Review, 2024, 12 (1)
- Accès au texte intégral et bibtex
-
- titre
- A self-regulatory cell-wall-sensing module at cell edges controls plant growth
- auteur
- Liam Elliott, Monika Kalde, Ann-Kathrin Schürholz, Xinyu Zhang, Sebastian Wolf, Ian Moore, Charlotte Kirchhelle
- article
- Nature Plants, In press, 10 (3), ⟨10.1038/s41477-024-01629-8⟩
- Accès au texte intégral et bibtex
-
- titre
- Water fluxes pattern growth and identity in shoot meristems
- auteur
- Juan Alonso-Serra, Ibrahim Cheddadi, Annamaria Kiss, Guillaume Cerutti, Marianne Lang, Sana Dieudonné, Claire Lionnet, Christophe Godin, Olivier Hamant
- article
- Nature Communications, 2024, 15 (1), pp.6944. ⟨10.1038/s41467-024-51099-x⟩
- Accès au texte intégral et bibtex
-
Preprints, Working Papers, ...
- titre
- Northern Normal – Laboratory Networks, Microbial Culture Collections, and Taxonomies of Power (1939-2000)
- auteur
- Claas Kirchhelle, Charlotte Kirchhelle
- article
- 2024
- Accès au bibtex
-
- titre
- Puzzle cell shape emerges from the interaction of growth with mechanical constraints
- auteur
- Nicola Trozzi, Brendan Lane, Alice Perruchoud, Yixuan Wang, Lukas Hörmayer, Mylan Ansel, Corentin Mollier, Alice Malivert, Frances Clark, Tammo Reichgelt, Adrienne H.K. Roeder, Olivier Hamant, Arezki Boudaoud, Dorota Kwiatkowska, Adam Runions, Richard Smith, Mateusz Majda
- article
- 2024
- Accès au bibtex
-
2023
Journal articles
- titre
- The plasma membrane – cell wall nexus in plant cells: focus on the Hechtian structure
- auteur
- Denise S Arico, Johanna E M Dickmann, Olivier Hamant, Hervé Canut
- article
- The Cell Surface, 2023, 10, pp.100115. ⟨10.1016/j.tcsw.2023.100115⟩
- Accès au texte intégral et bibtex
-
- titre
- Increased gene expression variability hinders the formation of regional mechanical conflicts leading to reduced organ shape robustness
- auteur
- Duy-Chi Trinh, Marjolaine Martin, Lotte Bald, Alexis Maizel, Christophe Trehin, Olivier Hamant
- article
- Proceedings of the National Academy of Sciences of the United States of America, 2023, 120 (30), pp.e2302441120. ⟨10.1073/pnas.2302441120⟩
- Accès au bibtex
-
- titre
- A numerical framework coupling finite element and meshless methods in sequential and parallel simulations
- auteur
- Van Dung Nguyen, Charlotte Kirchhelle, Amir Abdollahi, Julián Andrés García Grajales, Dongli Li, Kamel Benatia, Khariton Gorbunov, Sylvin Bielle, Alain Goriely, Antoine Jérusalem
- article
- Finite Elements in Analysis and Design, 2023, 219, pp.103927. ⟨10.1016/j.finel.2023.103927⟩
- Accès au texte intégral et bibtex
-
- titre
- Why is FERONIA pleiotropic?
- auteur
- Alice Malivert, Olivier Hamant
- article
- Nature Plants, 2023, 9 (7), pp.1018-1025. ⟨10.1038/s41477-023-01434-9⟩
- Accès au bibtex
-
- titre
- Discretizing the cellular bases of plant morphogenesis: Emerging properties from subcellular and noisy patterning
- auteur
- Charlotte Kirchhelle, Olivier Hamant
- article
- Current Opinion in Cell Biology, 2023, 81, pp.102159. ⟨10.1016/j.ceb.2023.102159⟩
- Accès au texte intégral et bibtex
-
- titre
- Contribution of vasculature to stem integrity in Arabidopsis thaliana
- auteur
- Mariko Asaoka, Mariko Asaoka, Shingo Sakamoto, Shizuka Gunji, Nobutaka Mitsuda, Hirokazu Tsukaya, Shinichiro Sawa, Olivier Hamant, Ali Ferjani
- article
- Development (Cambridge, England), 2023, 150 (3), ⟨10.1242/dev.201156⟩
- Accès au texte intégral et bibtex
-
- titre
- The 1972 Meadows report: A wake-up call for plant science
- auteur
- Olivier Hamant
- article
- Quantitative Plant Biology, 2023, 4, pp.e3. ⟨10.1017/qpb.2023.2⟩
- Accès au texte intégral et bibtex
-
Book sections
- titre
- Single-Cell Confinement Methods to Study Plant Cytoskeleton
- auteur
- Pauline Durand-Smet, Antoine Chevallier, Léia Colin, Alice Malivert, Isaty Melogno, Olivier Hamant
- article
- The Plant Cytoskeleton, 2604, Springer US; Springer US, pp.63-75, 2023, Methods in Molecular Biology, ⟨10.1007/978-1-0716-2867-6_5⟩
- Accès au bibtex
-
2022
Journal articles
- titre
- Endoreplication mediates cell size control via mechanochemical signaling from cell wall
- auteur
- Yuan Ma, Kristoffer Jonsson, Bibek Aryal, Lieven de Veylder, Olivier Hamant, Rishikesh Bhalerao
- article
- Science Advances , 2022, 8 (49), pp.eabq2047. ⟨10.1126/sciadv.abq2047⟩
- Accès au texte intégral et bibtex
-
- titre
- Shoot meristem progenitors emerge from mechanical heterogeneities
- auteur
- Olivier Hamant
- article
- Developmental Cell, 2022, 57 (17), pp.2043-2044. ⟨10.1016/j.devcel.2022.08.004⟩
- Accès au bibtex
-
- titre
- Looking beyond the gene network – metabolic and mechanical cell drivers of leaf morphogenesis
- auteur
- Hokuto Nakayama, Hiroyuki Koga, Yuchen Long, Olivier Hamant, Ali Ferjani
- article
- Journal of Cell Science, 2022, 135 (8), pp.jcs259611. ⟨10.1242/jcs.259611⟩
- Accès au bibtex
-
- titre
- Plant cell walls as mechanical signaling hubs for morphogenesis
- auteur
- Kristoffer Jonsson, Olivier Hamant, Rishikesh Bhalerao
- article
- Current Biology, 2022, 32 (7), pp.R334-R340. ⟨10.1016/j.cub.2022.02.036⟩
- Accès au texte intégral et bibtex
-
- titre
- Fifteen compelling open questions in plant cell biology
- auteur
- Adrienne H K Roeder, Marisa S Otegui, Ram Dixit, Charles T Anderson, Christine Faulkner, Yan Zhang, Maria J Harrison, Charlotte Kirchhelle, Gohta Goshima, Jeremy E Coate, Jeff J Doyle, Olivier Hamant, Keiko Sugimoto, Liam Dolan, Heather Meyer, David W Ehrhardt, Arezki Boudaoud, Carlos Messina
- article
- The Plant cell, 2022, 34 (1), pp.72-102. ⟨10.1093/plcell/koab225⟩
- Accès au texte intégral et bibtex
-
2021
Journal articles
- titre
- FERONIA and microtubules independently contribute to mechanical integrity in the Arabidopsis shoot
- auteur
- Alice Malivert, Özer Erguvan, Antoine Chevallier, Antoine Dehem, Rodrigue Friaud, Mengying Liu, Marjolaine Martin, Théophile Peyraud, Olivier Hamant, Stéphane Verger
- article
- PLoS Biology, 2021, 19 (11), pp.e3001454. ⟨10.1371/journal.pbio.3001454⟩
- Accès au bibtex
-
- titre
- Inducible depletion of PI(4,5)P2 by the synthetic iDePP system in Arabidopsis
- auteur
- Mehdi Doumane, Alexis Lebecq, Léia Colin, Aurélie Fangain, Floris Stevens, Joseph Bareille, Olivier Hamant, Youssef Belkhadir, Teun Munnik, Yvon Jaillais, Marie-Cécile Caillaud
- article
- Nature Plants, 2021, 7 (5), pp.587-597. ⟨10.1038/s41477-021-00907-z⟩
- Accès au texte intégral et bibtex
-
- titre
- Fluctuations shape plants through proprioception
- auteur
- Bruno Moulia, Stéphane Douady, Olivier Hamant
- article
- Science, 2021, 372 (6540), pp.eabc6868. ⟨10.1126/science.abc6868⟩
- Accès au bibtex
-
- titre
- Mechanochemical feedback mediates tissue bending required for seedling emergence
- auteur
- Kristoffer Jonsson, Rahul Lathe, Daniel Kierzkowski, Anne-Lise Routier-Kierzkowska, Olivier Hamant, Rishikesh Bhalerao
- article
- Current Biology, 2021, 31 (6), pp.1154-1164.e3. ⟨10.1016/j.cub.2020.12.016⟩
- Accès au texte intégral et bibtex
-
- titre
- Tissue folding at the organ–meristem boundary results in nuclear compression and chromatin compaction
- auteur
- Kateryna Fal, Niklas Korsbo, Juan Alonso-Serra, Jose Teles, Mengying Liu, Yassin Refahi, Marie-Edith Chaboute, Henrik Jönsson, Olivier Hamant
- article
- Proceedings of the National Academy of Sciences of the United States of America, 2021, 118 (8), pp.e2017859118. ⟨10.1073/pnas.2017859118⟩
- Accès au texte intégral et bibtex
-
- titre
- Stem integrity in Arabidopsis thaliana requires a load-bearing epidermis
- auteur
- Mariko Asaoka, Mao Ooe, Shizuka Gunji, Pascale Milani, Gaël Runel, Gorou Horiguchi, Olivier Hamant, Shinichiro Sawa, Hirokazu Tsukaya, Ali Ferjani
- article
- Development (Cambridge, England), 2021, 148 (4), ⟨10.1242/dev.198028⟩
- Accès au bibtex
-
- titre
- Plants Show Us the Light
- auteur
- Olivier Hamant
- article
- Trends in Plant Science, 2021, 26 (2), pp.97-99. ⟨10.1016/j.tplants.2020.10.011⟩
- Accès au texte intégral et bibtex
-
- titre
- How Mechanical Forces Shape Plant Organs
- auteur
- Duy-Chi Trinh, Juan Alonso-Serra, Mariko Asaoka, Leia Colin, Matthieu Cortes, Alice Malivert, Shogo Takatani, Feng Zhao, Jan Traas, Christophe Trehin, Olivier Hamant
- article
- Current Biology, 2021, 31 (3), pp.R143-R159. ⟨10.1016/j.cub.2020.12.001⟩
- Accès au texte intégral et bibtex
-
- titre
- External Mechanical Cues Reveal a Katanin-Independent Mechanism behind Auxin-Mediated Tissue Bending in Plants
- auteur
- Anirban Baral, Bibek Aryal, Kristoffer Jonsson, Emily Morris, Elsa Demes, Shogo Takatani, Stéphane Verger, Tongda Xu, Malcolm Bennett, Olivier Hamant, Rishikesh Bhalerao
- article
- Developmental Cell, 2021, 56 (1), pp.67-80.e3. ⟨10.1016/j.devcel.2020.12.008⟩
- Accès au bibtex
-
- titre
- The plasma membrane as a mechanotransducer in plants
- auteur
- Leia Colin, Olivier Hamant
- article
- Comptes Rendus. Biologies, 2021, 344 (4), pp.389-407. ⟨10.5802/crbiol.66⟩
- Accès au texte intégral et bibtex
-
- titre
- Organ geometry channels reproductive cell fate in the Arabidopsis ovule primordium
- auteur
- Elvira Hernandez-Lagana, Gabriella Mosca, Ethel Mendocilla-Sato, Nuno Pires, Anja Frey, Alejandro Giraldo-Fonseca, Caroline Michaud, Ueli Grossniklaus, Olivier Hamant, Christophe Godin, Arezki Boudaoud, Daniel Grimanelli, Daphné Autran, Célia Baroux
- article
- eLife, 2021, 10, pp.e66031. ⟨10.7554/eLife.66031⟩
- Accès au texte intégral et bibtex
-
- titre
- What is quantitative plant biology?
- auteur
- Daphné Autran, George W Bassel, Eunyoung Chae, Daphne Ezer, Ali Ferjani, Christian Fleck, Olivier Hamant, Félix Hartmann, Yuling Jiao, Iain G Johnston, Dorota Kwiatkowska, Boon L Lim, Ari Pekka Mahönen, Richard J Morris, Bela M Mulder, Naomi Nakayama, Ross Sozzani, Lucia C Strader, Kirsten ten Tusscher, Minako Ueda, Sebastian Wolf
- article
- Quantitative Plant Biology, 2021, 2, 16 p. ⟨10.1017/qpb.2021.8⟩
- Accès au texte intégral et bibtex
-
2020
Journal articles
- titre
- Microtubule self-organisation during seed germination in Arabidopsis
- auteur
- Huifang Yan, Nicole Chaumont, Jean François Gilles, Susanne Bolte, Olivier Hamant, Christophe Bailly
- article
- BMC Biology, 2020, 18 (1), ⟨10.1186/s12915-020-00774-8⟩
- Accès au texte intégral et bibtex
-
- titre
- Plant scientists can’t ignore Jevons paradox anymore
- auteur
- Olivier Hamant
- article
- Nature Plants, 2020, 6 (7), pp.720-722. ⟨10.1038/s41477-020-0722-3⟩
- Accès au bibtex
-
- titre
- How humans may co-exist with Earth? The case for suboptimal systems
- auteur
- Stéphane Grumbach, Olivier Hamant
- article
- Anthropocene, 2020, 30, pp.1-10. ⟨10.1016/j.ancene.2020.100245⟩
- Accès au texte intégral et bibtex
-
- titre
- Robust organ size requires robust timing of initiation orchestrated by focused auxin and cytokinin signalling
- auteur
- Mingyuan Zhu, Weiwei Chen, Vincent Mirabet, Lilan Hong, Simone Bovio, Soeren Strauss, Erich Schwarz, Satoru Tsugawa, Zhou Wang, Richard Smith, Chun-Biu Li, Olivier Hamant, Arezki Boudaoud, Adrienne Roeder
- article
- Nature Plants, 2020, 6 (6), pp.686-698. ⟨10.1038/s41477-020-0666-7⟩
- Accès au texte intégral et bibtex
-
- titre
- Microtubule Response to Tensile Stress Is Curbed by NEK6 to Buffer Growth Variation in the Arabidopsis Hypocotyl
- auteur
- Shogo Takatani, Stéphane Verger, Takashi Okamoto, Taku Takahashi, Olivier Hamant, Hiroyasu Motose
- article
- Current Biology, 2020, 30 (8), pp.1491-1503.e2. ⟨10.1016/j.cub.2020.02.024⟩
- Accès au texte intégral et bibtex
-
- titre
- Flowering Plants in the Anthropocene: A Political Agenda
- auteur
- Ioan Negrutiu, Michael Frohlich, Olivier Hamant
- article
- Trends in Plant Science, 2020, 25 (4), pp.349-368. ⟨10.1016/j.tplants.2019.12.008⟩
- Accès au bibtex
-
- titre
- Mechanical Shielding in Plant Nuclei
- auteur
- Rituparna Goswami, Atef Asnacios, Pascale Milani, Stéfanie Graindorge, Guy Houlné, Jérôme Mutterer, Olivier Hamant, Marie-Edith Chabouté
- article
- Current Biology, 2020, 30 (11), pp.2013-+. ⟨10.1016/j.cub.2020.03.059⟩
- Accès au texte intégral et bibtex
-
- titre
- Editorial
- auteur
- Olivier Hamant
- article
- Quantitative Plant Biology, 2020, 1, pp.e1. ⟨10.1017/qpb.2020.2⟩
- Accès au texte intégral et bibtex
-
- titre
- Is the plant nucleus a mechanical rheostat?
- auteur
- Rituparna Goswami, Atef Asnacios, Olivier Hamant, Marie-Edith Chabouté
- article
- Current Opinion in Plant Biology, 2020, 57, pp.155-163. ⟨10.1016/j.pbi.2020.09.001⟩
- Accès au texte intégral et bibtex
-
- titre
- Cortical tension overrides geometrical cues to orient microtubules in confined protoplasts
- auteur
- Leia Colin, Antoine Chevallier, Satoru Tsugawa, Florian Gacon, Christophe Godin, Virgile Viasnoff, Timothy Saunders, Olivier Hamant
- article
- Proceedings of the National Academy of Sciences of the United States of America, 2020, 117 (51), pp.32731-32738. ⟨10.1073/pnas.2008895117⟩
- Accès au texte intégral et bibtex
-
- titre
- Yves Couder: Putting mechanics back into the shoot apical meristem
- auteur
- Jan Traas, Olivier Hamant
- article
- Comptes Rendus. Mécanique, 2020, 348 (6-7), pp.679-684. ⟨10.5802/crmeca.19⟩
- Accès au bibtex
-
Book sections
- titre
- Shaping Organs: Shared Structural Principles Across Kingdoms
- auteur
- T.E. Saunders, Olivier Hamant
- article
- Annual Review of Cell and Developmental Biology, 36 (1), pp.385-410, 2020, ⟨10.1146/annurev-cellbio-012820-103850⟩
- Accès au bibtex
-
Preprints, Working Papers, ...
- titre
- iDePP: a genetically encoded system for the inducible depletion of PI(4,5)P 2 in Arabidopsis thaliana
- auteur
- Mehdi Doumane, Léia Colin, Alexis Lebecq, Aurélie Fangain, Joseph Bareille, Olivier Hamant, Youssef Belkhadir, Yvon Jaillais, Marie-Cécile Caillaud
- article
- 2020
- Accès au texte intégral et bibtex
-
2019
Journal articles
- titre
- Are microtubules tension sensors?
- auteur
- Olivier O. Hamant, Daisuke Inoue, David Bouchez, Jacques Dumais, Eric Mjolsness
- article
- Nature Communications, 2019, 10 (1), ⟨10.1038/s41467-019-10207-y⟩
- Accès au texte intégral et bibtex
-
- titre
- Does resource availability help determine the evolutionary route to multicellularity?
- auteur
- Olivier Hamant, Ramray Bhat, Vidyanand Nanjundiah, Stuart A. Newman
- article
- Evolution and Development, 2019, 21 (3), pp.115-119. ⟨10.1111/ede.12287⟩
- Accès au texte intégral et bibtex
-
- titre
- Mechanical asymmetry of the cell wall predicts changes in pavement cell geometry
- auteur
- Mateusz Majda, Pawel Krupinski, Henrik Jönsson, Olivier O. Hamant, Stéphanie Robert
- article
- Developmental Cell, 2019, 50 (1), pp.9-10. ⟨10.1016/j.devcel.2019.06.002⟩
- Accès au bibtex
-
- titre
- Mechanical conflicts in twisting growth revealed by cell-cell adhesion defects
- auteur
- Stéphane Verger, Mengying Liu, Olivier Hamant
- article
- Frontiers in Plant Science, 2019, 10, ⟨10.3389/fpls.2019.00173⟩
- Accès au texte intégral et bibtex
-
- titre
- Seven steps to make travel to scientific conferences more sustainable
- auteur
- Olivier Hamant, Timothy Saunders, Virgile Viasnoff
- article
- Nature, 2019, 573 (7774), pp.451-452. ⟨10.1038/d41586-019-02747-6⟩
- Accès au texte intégral et bibtex
-
- titre
- ImageJ SurfCut: a user-friendly pipeline for high-throughput extraction of cell contours from 3D image stacks
- auteur
- Ozer Erguvan, Marion Louveaux, Olivier Hamant, Stéphane Verger
- article
- BMC Biology, 2019, 17, ⟨10.1186/s12915-019-0657-1⟩
- Accès au texte intégral et bibtex
-
Book sections
- titre
- Time-Lapse Imaging of Developing Shoot Meristems Using A Confocal Laser Scanning Microscope
- auteur
- Olivier O. Hamant, Pradeep Das, Agata Burian
- article
- Plant Cell Morphogenesis: Methods and Protocols, 1992, pp.257-268, 2019, 978-1-4939-9469-4, 978-1-4939-9468-7. ⟨10.1007/978-1-4939-9469-4_17⟩
- Accès au bibtex
-
2018
Journal articles
- titre
- The contribution of mechanosensing to epidermal cell fate specification
- auteur
- Alice Malivert, Olivier Hamant, Gwyneth Ingram
- article
- Current Opinion in Genetics and Development, 2018, 51, pp.52-58. ⟨10.1016/j.gde.2018.06.011⟩
- Accès au bibtex
-
- titre
- An image analysis pipeline to quantify emerging cracks in materials or adhesion defects in living tissues
- auteur
- Stéphane Verger, Guillaume Cerutti, Olivier Hamant
- article
- Bio-protocol , 2018, 8 (19), pp.1-16. ⟨10.21769/BioProtoc.3036⟩
- Accès au texte intégral et bibtex
-
- titre
- A tension-adhesion feedback loop in plant epidermis
- auteur
- Stéphane Verger, Yuchen Long, Arezki Boudaoud, Olivier O. Hamant
- article
- eLife, 2018, 7, ⟨10.7554/elife.34460.001⟩
- Accès au texte intégral et bibtex
-
- titre
- A phosphoinositide map at the shoot apical meristem in Arabidopsis thaliana
- auteur
- Thomas Stanislas, Matthieu Platre, Mengying Liu, Léa E S Rambaud-Lavigne, Yvon Jaillais, Olivier Hamant
- article
- BMC Biology, 2018, 16 (1), pp.20. ⟨10.1186/s12915-018-0490-y⟩
- Accès au texte intégral et bibtex
-
- titre
- The self-organization of plant microtubules inside the cell volume yields their cortical localization, stable alignment, and sensitivity to external cues
- auteur
- Vincent Mirabet, Pawel Krupinski, Olivier O. Hamant, Elliot M. Meyerowitz, Henrik Jönsson, Arezki Boudaoud
- article
- PLoS Computational Biology, 2018, 14 (2), pp.e1006011. ⟨10.1371/journal.pcbi.1006011⟩
- Accès au texte intégral et bibtex
-
- titre
- Plant physiology: FERONIA defends the cell walls against corrosion
- auteur
- Stéphane Verger, Olivier O. Hamant
- article
- Current Biology, 2018, 28 (5), pp.R215-R217. ⟨10.1016/j.cub.2018.01.043⟩
- Accès au bibtex
-
- titre
- Why plants make puzzle cells, and how their shape emerges
- auteur
- Aleksandra Sapala, Adam Runions, Anne-Lise Routier-Kierzkowska, Mainak das Gupta, Lilan Hong, Hugo Hofhuis, Stéphane Verger, Gabriella Mosca, Chun-Biu Li, Angela Hay, Olivier O. Hamant, Adrienne H. K. Roeder, Miltos Tsiantis, Przemyslaw Prusinkiewicz, Richard S. Smith
- article
- eLife, 2018, 7, ⟨10.7554/elife.32794⟩
- Accès au texte intégral et bibtex
-
- titre
- An Image Analysis Pipeline to Quantify Emerging Cracks in Materials or Adhesion Defects in Living Tissues
- auteur
- Stéphane Verger, Guillaume Cerutti, Olivier Hamant
- article
- Bio-protocol , 2018, 8 (19)
- Accès au texte intégral et bibtex
-
- titre
- Heterogeneity and robustness in plant morphogenesis: From cells to organs
- auteur
- Lilan Hong, Mathilde Dumond, Mingyuan Zhu, Satoru Tsugawa, Chun-Biu Li, Arezki Boudaoud, Olivier O. Hamant, Adrienne H.K. Roeder
- article
- Annual Review of Plant Biology, 2018, 69 (1), pp.469-495. ⟨10.1146/annurev-arplant-042817-040517⟩
- Accès au bibtex
-
2017
Journal articles
- titre
- Digital Revolution or Anthropocenic Feedback?
- auteur
- Stéphane Grumbach, Olivier Hamant
- article
- Anthropocene Review, 2017, 5 (1), pp.1-12. ⟨10.1177/2053019617748337⟩
- Accès au texte intégral et bibtex
-
- titre
- Life behind the wall: sensing mechanical cues in plants
- auteur
- Olivier Hamant, Elizabeth S. Haswell
- article
- BMC Biology, 2017, 15 (1), pp.1-9. ⟨10.1186/s12915-017-0403-5⟩
- Accès au texte intégral et bibtex
-
- titre
- Nuclear envelope : a new frontier in plant mechanosensing ?
- auteur
- Kateryna Fal, A Asnacios, Marie-Edith Chaboute, Olivier Hamant
- article
- Biophysical Reviews, 2017, 9 (4), pp.389-403. ⟨10.1007/s12551-017-0302-6⟩
- Accès au texte intégral et bibtex
-
- titre
- Pourquoi "maintenant" ? Le cas de la révolution de l’intermédiation algorithmique
- auteur
- Stéphane Grumbach, Olivier O. Hamant
- article
- Multitudes, A paraître
- Accès au bibtex
-
- titre
- The RNA polymerase-associated factor 1 complex Is required for plant touch responses
- auteur
- Gregory S. Jensen, Kateryna Fal, Olivier Hamant, Elizabeth S. Haswell
- article
- Journal of Experimental Botany, 2017, 68 (3), pp.499-511. ⟨10.1093/jxb/erw439⟩
- Accès au texte intégral et bibtex
-
- titre
- Mechanochemical polarization of contiguous cell walls shapes plant pavement cells
- auteur
- Mateusz Majda, Peter Grones, Ida-Maria Sintorn, Thomas Vain, Pascale Milani, Pawel Krupinski, Beata Zagórska-Marek, Corrado Viotti, Henrik Jönsson, Ewa J. Mellerowicz, Olivier O. Hamant, Stéphanie Robert
- article
- Developmental Cell, 2017, 43 (3), pp.290-304. ⟨10.1016/j.devcel.2017.10.017⟩
- Accès au texte intégral et bibtex
-
- titre
- Mechano-devo
- auteur
- Olivier Hamant
- article
- Mechanisms of Development, 2017, 145, pp.2-9. ⟨10.1016/j.mod.2017.02.004⟩
- Accès au bibtex
-
- titre
- Mechanical shielding of rapidly growing cells buffers growth heterogeneity and contributes to organ shape reproducibility
- auteur
- Nathan Hervieux, Satoru Tsugawa, Antoine Fruleux, Mathilde Dumond, Anne-Lise Routier-Kierzkowska, Tamiki Komatsuzaki, Arezki Boudaoud, John C. Larkin, Richard S. Smith, Chun-Biu Li, Olivier O. Hamant
- article
- Current Biology, 2017, 27 (22), pp.3468-3479. ⟨10.1016/j.cub.2017.10.033⟩
- Accès au bibtex
-
- titre
- Clones of cells switch from reduction to enhancement of size variability in Arabidopsis sepals
- auteur
- Satoru Tsugawa, Nathan Hervieux, Daniel Kierzkowski, Anne-Lise Routier-Kierzkowska, Aleksandra Sapala, Olivier O. Hamant, Richard S. Smith, Adrienne H. K. Roeder, Arezki Boudaoud, Chun-Biu Li
- article
- Development (Cambridge, England), 2017, 144 (23), pp.4398-4405. ⟨10.1242/dev.153999⟩
- Accès au texte intégral et bibtex
-
- titre
- Phyllotactic regularity requires the Paf1 complex in Arabidopsis
- auteur
- Kateryna Fal, Mengying Liu, Assem Duisembekova, Yassin Refahi, Elizabeth S. Haswell, Olivier O. Hamant
- article
- Development (Cambridge, England), 2017, 144 (23), pp.4428-4436. ⟨10.1242/dev.154369⟩
- Accès au texte intégral et bibtex
-
Book sections
- titre
- In-vivo analysis of morphogenesis in plants
- auteur
- Thomas Stanislas, Olivier O. Hamant, Jan Traas
- article
- Cell Polarity and Morphogenesis, 139, Academic Press - Elsevier, 2017, Methods in Cell Biology, 978-0-12-809373-3. ⟨10.1016/bs.mcb.2016.11.008⟩
- Accès au bibtex
-
2016
Journal articles
- titre
- A mechanical feedback restricts sepal growth and shape in arabidopsis
- auteur
- Nathan Hervieux, Mathilde Dumond, Aleksandra Sapala, Anne-Lise Routier-Kierzkowska, Daniel Kierzkowski, Adrienne Roeder, Richard Smith, Arezki Boudaoud, Olivier O. Hamant
- article
- Current Biology, 2016, 26 (8), pp.1019-1028. ⟨10.1016/j.cub.2016.03.004⟩
- Accès au bibtex
-
- titre
- Variable cell growth yields reproducible organ development through spatiotemporal averaging
- auteur
- Lilan Hong, Mathilde Dumond, Satoru Tsugawa, Aleksandra Sapala, Anne-Lise Routier-Kierzkowska, Yong Zhou, Catherine Chen, Annamaria Kiss, Mingyuan Zhu, Olivier O. Hamant, Richard s. Smith, Tamiki Komatsuzaki, Chun-Biu Li, Arezki Boudaoud, Adrienne h.K. Roeder
- article
- Developmental Cell, 2016, 38 (1), pp.15-32. ⟨10.1016/j.devcel.2016.06.016⟩
- Accès au texte intégral et bibtex
-
- titre
- Cell division plane orientation based on tensile stress inArabidopsis thaliana
- auteur
- Marion Louveaux, Jean-Daniel Julien, Vincent Mirabet, Arezki Boudaoud, Arezki Boudaoud, Olivier O. Hamant
- article
- Proceedings of the National Academy of Sciences of the United States of America, 2016, 113 (30), pp.E4294-E4303. ⟨10.1073/pnas.1600677113⟩
- Accès au texte intégral et bibtex
-
- titre
- How do plants read their own shapes?
- auteur
- Olivier Hamant, Bruno Moulia
- article
- New Phytologist, 2016, 212 (2), pp.333-337. ⟨10.1111/nph.14143⟩
- Accès au bibtex
-
- titre
- Meristem Biology Flourishes Under Mt. Tai
- auteur
- Yuling Jiao, Olivier Hamant, Zhaojun Ding, Xian Sheng Zhang
- article
- Molecular Plant, 2016, 9 (8), pp.1224-1227. ⟨10.1016/j.molp.2016.07.001⟩
- Accès au bibtex
-
- titre
- Developing a 'thick skin': a paradoxical role for mechanical tension in maintaining epidermal integrity?
- auteur
- Roberta Galletti, Stéphane Verger, Olivier Hamant, Gwyneth Ingram
- article
- Development (Cambridge, England), 2016, 143 (18), pp.3249-3258. ⟨10.1242/dev.132837⟩
- Accès au texte intégral et bibtex
-
- titre
- An epidermis-driven mechanism positions and scales stem cell niches in plants
- auteur
- Jérémy Gruel, Benoit Landrein, Paul Tarr, Christoph Schuster, Yassin Refahi, Arun Sampathkumar, Olivier O. Hamant, Elliot M. Meyerowitz, Henrik Jönsson
- article
- Science Advances , 2016, 2 (1), ⟨10.1126/sciadv.1500989⟩
- Accès au texte intégral et bibtex
-
- titre
- Extracting subcellular fibrillar alignment with error estimation: Application to microtubules
- auteur
- Satoru Tsugawa, Nathan Hervieux, Olivier O. Hamant, Arezki Boudaoud, Richard S. Smith, Chun-Biu Li, Tamiki Komatsuzaki
- article
- Biophysical Journal, 2016, 110 (8), pp.1836-1844. ⟨10.1016/j.bpj.2016.03.011⟩
- Accès au texte intégral et bibtex
-
- titre
- The impact of mechanical compression on cortical microtubules in Arabidopsis: a quantitative pipeline
- auteur
- Marion Louveaux, Sebastien Rochette, Léna Beauzamy, Arezki Boudaoud, Olivier Hamant
- article
- The Plant Journal, 2016, 88 (2), pp.328-342. ⟨10.1111/tpj.13290⟩
- Accès au texte intégral et bibtex
-
- titre
- Interplay between miRNA regulation and mechanical stress for CUC gene expression at the shoot apical meristem
- auteur
- Kateryna Fal, Benoit Landrein, Olivier O. Hamant
- article
- Plant Signaling and Behavior, 2016, 11 (3), ⟨10.1080/15592324.2015.1127497⟩
- Accès au bibtex
-
2015
Journal articles
- titre
- A Computational Framework for 3D Mechanical Modeling of Plant Morphogenesis with Cellular Resolution
- auteur
- Frédéric Boudon, Jérôme Chopard, Olivier Ali, Benjamin Gilles, Olivier Hamant, Arezki Boudaoud, Jan Traas, Christophe Godin
- article
- PLoS Computational Biology, 2015, 11 (1), pp.1-16. ⟨10.1371/journal.pcbi.1003950⟩
- Accès au texte intégral et bibtex
-
- titre
- Meristem size contributes to the robustness of phyllotaxis in Arabidopsis
- auteur
- Benoit Landrein, Yassin Refahi, Fabrice Besnard, Nathan Hervieux, Vincent Mirabet, Arezki Boudaoud, Teva Vernoux, Olivier O. Hamant
- article
- Journal of Experimental Botany, 2015, 66 (5), pp.1317-1324. ⟨10.1093/jxb/eru482⟩
- Accès au texte intégral et bibtex
-
- titre
- Mechanical stress contributes to the expression of the STM homeobox gene in Arabidopsis shoot meristems
- auteur
- Benoit Landrein, Annamaria Kiss, Massimiliano Sassi, Aurélie Chauvet, Pradeep Das, Millan Cortizo Sabugo Cortizo, Patrick P. Laufs, Seiji Takeda, Mitsuhiro Aida, Jan Traas, Teva Vernoux, Arezki Boudaoud, Olivier O. Hamant
- article
- eLife, 2015, 4, pp.1-27. ⟨10.7554/eLife.07811⟩
- Accès au texte intégral et bibtex
-
- titre
- Mechanically, the shoot apical meristem of Arabidopsis behaves like a shell inflated by a pressure of about 1 MPa
- auteur
- Léna Beauzamy, Marion Louveaux, Olivier O. Hamant, Arezki Boudaoud
- article
- Frontiers in Plant Science, 2015, 6, pp.1-10. ⟨10.3389/fpls.2015.01038⟩
- Accès au texte intégral et bibtex
-
2014
Journal articles
- titre
- An Auxin-Mediated Shift toward Growth Isotropy Promotes Organ Formation at the Shoot Meristem in Arabidopsis
- auteur
- Massimiliano Sassi, Olivier Ali, Frédéric Boudon, Gladys Cloarec, Ursula Abad, Coralie Cellier, Xu Chen, Benjamin Gilles, Pascale Milani, Jìrí Friml, Teva Vernoux, Christophe Godin, Olivier Hamant, Jan Traas
- article
- Current Biology, 2014, 24 (19), pp.2335-2342. ⟨10.1016/j.cub.2014.08.036⟩
- Accès au texte intégral et bibtex
-
- titre
- FibrilTool, an ImageJ plug-in to quantify fibrillar structures in raw microscopy images
- auteur
- Arezki Boudaoud, Agata Burian, Dorota Borowska-Wykret, Magalie Uyttewaal, Roman Wrzalik, Dorota Kwiatkowska, Olivier Hamant
- article
- Nature Protocols, 2014, 9 (2), pp.457 - 463. ⟨10.1038/nprot.2014.024⟩
- Accès au bibtex
-
- titre
- A Comparative Mechanical Analysis of Plant and Animal Cells Reveals Convergence across Kingdoms
- auteur
- Pauline Durand-Smet, Nicolas Chastrette, Axel Guiroy, Alain Richert, Annick Berne-Dedieu, Judit Szécsi, Arezki Boudaoud, Jean-Marie Frachisse, Mohammed Bendahmane, Olivier O. Hamant, Atef Asnacios
- article
- Biophysical Journal, 2014, 107 (10), pp.2237-2244. ⟨10.1016/j.bpj.2014.10.023⟩
- Accès au bibtex
-
- titre
- Matching Patterns of Gene Expression to Mechanical Stiffness at Cell Resolution through Quantitative Tandem Epifluorescence and Nanoindentation.
- auteur
- Pascale Milani, Vincent Mirabet, Coralie Cellier, Frédérique Rozier, Olivier O. Hamant, Pradeep Das, Arezki Boudaoud
- article
- Plant Physiology, 2014, 165 (4), pp.1399-1408. ⟨10.1104/pp.114.237115⟩
- Accès au texte intégral et bibtex
-
- titre
- Subcellular and supracellular mechanical stress prescribes cytoskeleton behavior in Arabidopsis cotyledon pavement cells
- auteur
- Arun Sampathkumar, Pawel Krupinski, Raymond Wightman, Pascale Milani, Alexandre Berquand, Arezki Boudaoud, Olivier O. Hamant, Henrik Jönsson, Elliot M. Meyerowitz
- article
- eLife, 2014, 3, ⟨10.7554/eLife.01967.001⟩
- Accès au texte intégral et bibtex
-
Book sections
- titre
- Time-lapse imaging of developing meristems using confocal laser scanning microscope
- auteur
- Olivier O. Hamant, Pradeep Das, Agata Burian
- article
- Plant cell morphogenesis: methods and protocols, 1080, Humana Press, 2014, Methods in Molecular Biology, 978-1-62703-643-6; 978-1-62703-642-9. ⟨10.1007/978-1-62703-643-6_9⟩
- Accès au bibtex
-
2013
Journal articles
- titre
- Integrative cell biology: katanin at the crossroads
- auteur
- Olivier O. Hamant
- article
- Current Biology, 2013, 23 (5), pp.R206-R208. ⟨10.1016/j.cub.2013.01.031⟩
- Accès au bibtex
-
- titre
- Plant science and agricultural productivity: Why are we hitting the yield ceiling?
- auteur
- Steve de Bossoreille de Ribou, Florian Douam, Olivier O. Hamant, Michael W. Frohlich, Ioan Negrutiu
- article
- Plant Science, 2013, 210, pp.159-176. ⟨10.1016/j.plantsci.2013.05.010⟩
- Accès au bibtex
-
- titre
- How mechanical stress controls microtubule behavior and morphogenesis in plants: history, experiments and revisited theories
- auteur
- Benoit Landrein, Olivier O. Hamant
- article
- The Plant Journal, 2013, 75 (2), pp.324 - 338. ⟨10.1111/tpj.12188⟩
- Accès au bibtex
-
- titre
- A correlative microscopy approach relates microtubule behaviour, local organ geometry, and cell growth at the Arabidopsis shoot apical meristem
- auteur
- Agata Burian, Michal Ludynia, Magalie Uyttewaal, Jan Traas, Arezki Boudaoud, Olivier Hamant, Dorota Kwiatkowska
- article
- Journal of Experimental Botany, 2013, 64 (18), pp.5753-5767. ⟨10.1093/jxb/ert352⟩
- Accès au texte intégral et bibtex
-
- titre
- Cell biology: cytoskeleton network topology feeds back on its regulation
- auteur
- Olivier O. Hamant
- article
- Current Biology, 2013, 23 (21), pp.R963-R965. ⟨10.1016/j.cub.2013.09.051⟩
- Accès au bibtex
-
- titre
- Widespread mechanosensing controls the structure behind the architecture in plants.
- auteur
- Olivier O. Hamant
- article
- Current Opinion in Plant Biology, 2013, 16 (5), pp.654-60. ⟨10.1016/j.pbi.2013.06.006⟩
- Accès au bibtex
-
- titre
- Impaired Cellulose Synthase Guidance Leads to Stem Torsion and Twists Phyllotactic Patterns in Arabidopsis
- auteur
- Benoit Landrein, Rahul Lathe, Martin Bringmann, Cyril Vouillot, Alexander Ivakov, Arezki Boudaoud, Staffan Persson, Olivier O. Hamant
- article
- Current Biology, 2013, 23 (10), pp.895 - 900. ⟨10.1016/j.cub.2013.04.013⟩
- Accès au bibtex
-
- titre
- The mechanics behind cell division
- auteur
- Marion Louveaux, Olivier O. Hamant
- article
- Current Opinion in Plant Biology, 2013, 16 (6), pp.774-779. ⟨10.1016/j.pbi.2013.10.011⟩
- Accès au bibtex
-
2012
Journal articles
- titre
- Regulatory role of cell division rules on tissue growth heterogeneity
- auteur
- Karen Alim, Olivier O. Hamant, Arezki Boudaoud
- article
- Frontiers in Plant Science, 2012, 3, pp.1-13. ⟨10.3389/fpls.2012.00174⟩
- Accès au texte intégral et bibtex
-
- titre
- Cracking the elusive alignment hypothesis: the microtubule-cellulose synthase nexus unraveled
- auteur
- Martin Bringmann, Benoit Landrein, Christian Schudoma, Olivier O. Hamant, Marie-Theres Hauser, Staffan Persson
- article
- Trends in Plant Science, 2012, 17 (11), pp.666 - 674. ⟨10.1016/j.tplants.2012.06.003⟩
- Accès au bibtex
-
- titre
- Mechanical Stress Acts via Katanin to Amplify Differences in Growth Rate between Adjacent Cells in Arabidopsis
- auteur
- Magalie M. Uyttewaal, Agata A. Burian, Karen K. Alim, Benoi T. B. T. Landrein, Dorota D. Borowska-Wykret, Annick A. Dedieu, Alexis A. Peaucelle, Michal M. Ludynia, Jan J. Traas, Arezki A. Boudaoud, Dorota D. Kwiatkowska, Olivier O. Hamant
- article
- Cell, 2012, 149 (2), pp.439 - 451. ⟨10.1016/j.cell.2012.02.048⟩
- Accès au bibtex
-
- titre
- The mechanics behind cell polarity
- auteur
- Atef Asnacios, Olivier O. Hamant
- article
- Trends in Cell Biology, 2012, 22 (11), pp.584 - 591. ⟨10.1016/j.tcb.2012.08.005⟩
- Accès au bibtex
-
2011
Journal articles
- titre
- Organogenesis from stem cells in planta: multiple feedback loops integrating molecular and mechanical signals
- auteur
- Fabrice Besnard, Teva Vernoux, Olivier O. Hamant
- article
- Cellular and Molecular Life Sciences, 2011, 68 (17), pp.2885-2906. ⟨10.1007/s00018-011-0732-4⟩
- Accès au bibtex
-
- titre
- The Role of Mechanical Forces in Plant Morphogenesis
- auteur
- Vincent Mirabet, Pradeep Das, Arezki Boudaoud, Olivier O. Hamant
- article
- Annual Review of Plant Biology, 2011, 62, pp.365 - 385. ⟨10.1146/annurev-arplant-042110-103852⟩
- Accès au bibtex
-
- titre
- Quantitative imaging strategies pave the way for testable biological concepts
- auteur
- Olivier O. Hamant
- article
- BMC Biology, 2011, 9, ⟨10.1186/1741-7007-9-10⟩
- Accès au texte intégral et bibtex
-
2010
Journal articles
- titre
- Integrating physical stress, growth and development.
- auteur
- M. Uyttewaal, Jan Traas, Olivier Hamant
- article
- Current Opinion in Plant Biology, 2010, 13 (1), pp.46-52
- Accès au bibtex
-
- titre
- Alignment between PIN1 polarity and microtubule orientation in the shoot apical meristem reveals a tight coupling between morphogenesis and auxin transport
- auteur
- Marcus G. Heisler, Olivier O. Hamant, Pawel Krupinski, Magalie M. Uyttewaal, Carolyn Ohno, Henrik Jönsson, Jan Traas, Elliot M. Meyerowitz
- article
- PLoS Biology, 2010, 8 (10), ⟨10.1371/journal.pbio.1000516⟩
- Accès au texte intégral et bibtex
-
- titre
- The mechanics behind plant development.
- auteur
- Olivier Hamant, Jan Traas
- article
- New Phytologist, 2010, 185 (2), pp.369-385. ⟨10.1111/j.1469-8137.2009.03100.x⟩
- Accès au bibtex
-
- titre
- Regulation of shape and patterning in plant development.
- auteur
- Olivier Hamant, Jan Traas, Arezki Boudaoud
- article
- Current Opinion in Genetics and Development, 2010, 20, pp.454-459
- Accès au bibtex
-
- titre
- Plant development: a TALE story
- auteur
- Olivier Hamant, Veronique Pautot
- article
- Comptes Rendus. Biologies, 2010, 333 (4), pp.371-381. ⟨10.1016/j.crvi.2010.01.015⟩
- Accès au bibtex
-
2009
Journal articles
- titre
- Turning a plant tissue into a living cell froth through isotropic growth
- auteur
- Francis Corson, Olivier Hamant, S. Bohn, Jan Traas, Arezki Boudaoud, Y. Couder
- article
- Proceedings of the National Academy of Sciences of the United States of America, 2009, 106 (21), pp.8453-8458
- Accès au bibtex
-
2008
Journal articles
- titre
- Developmental patterning by mechanical signals in Arabidopsis
- auteur
- Olivier Hamant, Mg Heisler, P. Krupinski, M. Uyttewaal, Francis Corson, Arezki Boudaoud, Em Meyerowitz, Y. Couder, P. Sahlin, Hannes Jónsson
- article
- Science, 2008, 322, pp.1650-1655
- Accès au bibtex
-
2006
Journal articles
- titre
- Alleles of add 1 dissect REC8 functions during meiotic prophase 1
- auteur
- Inna Golubovskaya, Olivier Hamant, L. Timofejeva, R. Wang Chung Ju, David Braun, Robert Meeley, Wz Cande
- article
- Journal of Cell Science, 2006, 119 (16), pp.3306-3315
- Accès au bibtex
-
- titre
- Genetics of meiotic prophase I in plants
- auteur
- Olivier Hamant, H.-L. Ma, Wz Cande
- article
- Annual Review of Plant Biology, 2006, 57, pp.267-302
- Accès au bibtex
-
- titre
- KNAT6: an Arabidopsis homeobox gene involved in meristem activity and organ separation.
- auteur
- E. Belles-Boix, Olivier Hamant, S.M. Witiak, H.L. Morin, Jan Traas, V. Pautot
- article
- The Plant cell, 2006, pp.1900-1907
- Accès au bibtex
-
Plant organ morphogenesis: geometry calls for biochemistry and mechanics
Changing shape is changing structure. By definition, this involves the laws of mechanics. Not only do plant generate diverse shapes, but shape itself can serve as an instructive cue to channel morphogenesis, through biochemical and mechanical signaling.
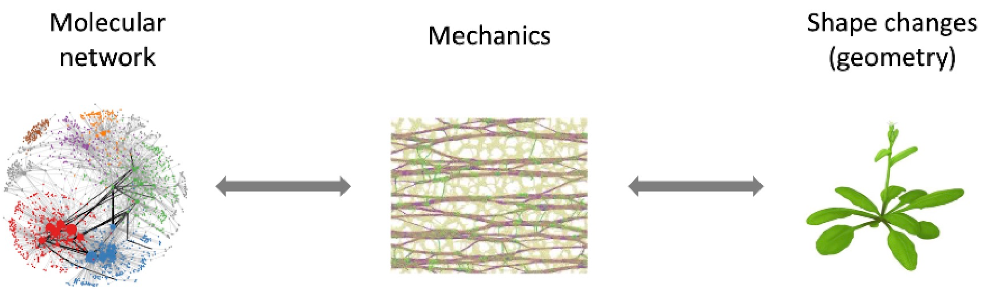
New scientific questions: robustness, multiscale hierarchy, emergent properties
How do organs know when to stop growing? How do multicellular organisms produce their diverse shapes? The underlying process, morphogenesis, is complex and characterised by three remarkable features: robustness, hierarchy, and emergence. Morphogenesis robustly produces highly similar forms across a wide range of conditions, despite and sometimes thanks to high levels of local variability. Morphogenesis involves events on a hierarchy of different scales in time and space (from the molecular to the organ scale). Individual factors contributing to morphogenetic events have been studied extensively, but are insufficient to fully predict morphogenesis – morphogenesis has emergent properties. Therefore, a central and challenging question in biology is how different factors are integrated and coordinated across multiple spatio-temporal scales to robustly produce organ shapes.
1. Cell Edges and Morphogenesis
Plants face a particular challenge when forming new organs: their cells share a rigid cell wall, which requires adjacent cells to coordinate their growth.

Growth patterns in Arabidopsis lateral roots
Directional growth depends on differences in cell wall mechanical properties in different regions of the cell. To establish and modulate these, cells precisely coordinate the transport of different cell wall components and their associated biosynthetic machinery to different regions of the cell surface.

Cell edges are a biochemically distinct domain in plant cells. Cell wall biosynthesis involves multiple trafficking pathways to the cell surface, which are regulated by Rab GTPases. RAB-A5c mediates a trafficking pathway specifically to cell edges in Arabidopsis lateral roots.
Our team is particularly interested in cell edges, the geometric domain at the intersection of two cell faces. Our previous work has shown that plants specify a transport route to cell edges which required for directional growth, but does not act through oriented cellulose deposition, the leading paradigm for directional growth control.
In our current work funded by ERC Starting Grant EDGE-CAM, we try to understand why cell edges are important during morphogenesis, and how plant cells specify their edges as distinct domains.
Why cell edges are important during morphogenesis? Cell edges are notable domains from a topological, geometric, and mechanical perspective. With respect to mechanics, they accumulate stresses arising at the cell level through turgor pressure, and are also exposed to shear stresses arising through differential growth at the tissue scale. We hypothesise that plants specify a cell wall sensing module at cell edges through which they can monitor the mechanical status at the cell wall and adapt growth through fine-tuning trafficking pathways. We are combining forward genetics, proteomics, quantitative imaging, and computational modelling to test our hypothesis and identify new components of edge-based growth control.
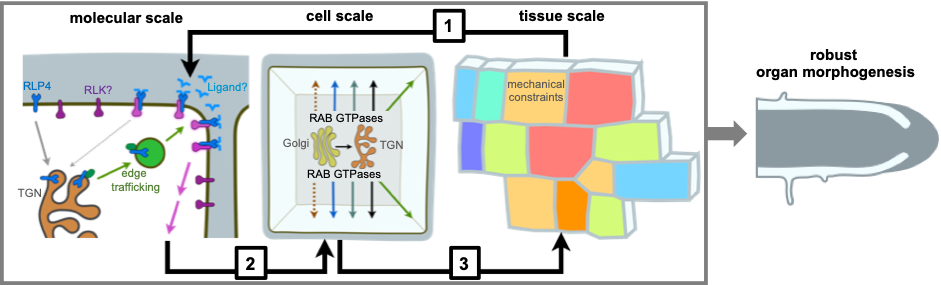
A multi-scale model for robust morphogenesis. [1] An RLP-based cell wall sensing module at cell edges senses cell wall changes associated with tissue-level mechanics. [2] Downstream targets of the cell wall sensing module include endomembrane trafficking pathways involved in cell wall assembly. [3] Activity of these pathways determines cellular growth patterns and tissue-level mechanical constraints.
How plant cells specify their edges as distinct domains? Protein localisation to cell edges has been linked to cytoskeleton organisation and the status of the cell wall, however it is an open question how plant cells identify their edges or differentiate between different edges in the same cell. We are exploring mechanical, geometric, and biochemical factors that may contribute to the recruitment of proteins to cell edges using quantitative imaging and genetics in planta and in an in vitro single-cell system.
Associated team members:
Charlotte Kirchhelle
Antoine Chevallier
Liam Elliott
Nathan German
Zoe Nemec Venza
Claire Lionnet
Marjolaine Martin
2. Multiscale microtubule response to stress

In past work, we showed that cortical microtubules align along maximal tensile stress directions, thereby reinforcing cell walls through the guidance of cellulose deposition.
Cortical microtubules also guide the orientation of the next division plane (through the preprophase band). Combining modeling and experiments, we formally showed that tensile stress prescribes cell division plane orientation. We also unraveled a role of microtubule dynamics and response to stress in organ initiation in organ shape robustness, in organ growth arrest and in organ flatness.

Confined protoplasts in microwells
We are now investigating the possibility that microtubules may be their own mechanosensors, notably using in vitro and microfabrication techniques, such as confined protoplasts in microwells.
In planta, this response may have important implications, relating mechanical conflicts due to differential growth to morphogenesis.

CreLox lines inducing growth mosaics and artificial mechanical conflicts at the shoot apex
Associated team members:
Olivier Hamant
Annalisa Bellandi
Antoine Chevallier
Charlotte Kirchhelle
Claire Lionnet
Marjolaine Martin
Isaty Melogno
Mariana Romeiro Motta
3. Mechanical identity
Beyond the cell cortex, we started to analyze the impact of mechanical signals on gene expression. We found that the expression of key transcription factors (CUC3 and STM) is in part under mechanical control.

STM expression in meristem boundaries, stress hotspots (Landrein et al., 2015)
The nexus between mechanical signals at the cell cortex and gene expression may involve different pathways. Recently, we started to analyze the role of nucleus deformation in gene expression.
We are also investighating the role of the RNA Polymerase-associated factor 1 complex (Paf1c), a central regulator of transcription, in development, at the nexus between transcriptional noise and developmental robustness. We found that Paf1c-dependent transcription contributes to the robustness of phyllotaxis and flower termination. Although at this stage it remains difficult to link mechanosensing with these phenotypes, these results open the possibility for crosstalks between transcriptional noise, mechanotransduction and development.

Severe indetermincay defects in the Paf1c mutant
Associated team members:
Olivier Hamant & Christophe Tréhin
Johanna Dickmann
Denise Arico
Marianne Lang
Claire Lionnet
Marjolaine Martin
4. Science and society
We are also involved in science-society projects, notably on the environmental question sustainability and in several art-science projects. This notably includes the question of robustness.
The case of pollard trees or trunks, of which all the upper and lateral branches are pruned, illustrates the remarkable plant plasticity. The trunks indeed develop a characteristic phenotype with an increased trunk thickness and massive repetitions of shoots. Since the Neolithic, this rural practice has been an essential source of renewable wood and fodder, before declining with the advent of the modern era. Conventional agriculture is increasingly under the spotlight for its negative externalities (greenhouse gas emissions,
pollution, dependence on fossil fuels, loss of biodiversity, desertification ...) and for its low resilience. From this perspective, the pollards find a new agronomic relevance.

Pollard tree drawing
Last, we are also investigating the potential use of plant cell walls as metamaterials, notably their phononic properties. There are numerous examples of animals that manipulate visible light for functional purposes, such as insect wings made of hexagon arrays of cone-shaped nanopillars that provide a graded refractive index for camouflage. Such photonic structures can be mimicked to find innovative solutions for light control in nano- to microstructures. In contrast, there has been no observation of phonon-based biological functions at the supramolecular scale, making phononic bioinspired strategies inoperative. Thomas Dehoux team has recently demonstrated that biological composites in the form of decellularized plant cell scaffolds can behave as phononic materials, including forbidding the propagation of elastic waves in select frequency ranges (i.e. band gaps). This opens the possibility for biobased phononic materials (BPM).

Propagation of an acoustic wave in an onion epidermal peel (T. Dehoux team)
Associated team members:
Olivier Hamant
Sana Dieudonné
Marianne Lang
To explore more science & society questions, visit the Michel Serres Institute website: https://institutmichelserres.fr/