






Publication du Laboratoire de géologie de Lyon : Terre, planètes et environnement (LGLTPE) dans la revue Scientific Reports du 28 août 2018.
Les oiseaux modernes n’ont pas de dents, mais un groupe éteint de grands oiseaux de mer, les odontopterygiformes, possédait des excroissances osseuses sur les mâchoires, ressemblant à des dents, et distribuées de façon séquentielle suivant leur taille. L’origine de ces "pseudodents" est restée jusqu’ici énigmatique. Sur la base de récents travaux et de nouveaux éléments, un modèle de développement de ces structures uniques et disparues est proposé. Une régulation développementale de type odontogénétique* a été détournée au cours de l’évolution et a conduit à une interaction entre épithélium* et cellules externes de l’os des mâchoires. Les ostéoblastes (les cellules permettant la formation des os) étaient capables de répondre à la signalisation épithéliale encore efficace chez ces oiseaux, à la place des odontoblastes absents (cellules qui interviennent dans la formation des dents). Des pseudodents croissaient sur les bords du bec à la place de vraies dents. Des "champs morphogénétiques" dynamiques agissaient au cours de la croissance des pseudodents, expliquant leur distribution unique le long des mâchoires en fonction de leur taille. Ainsi, le développement des pseudodents apparaît comme une homologie* profonde avec le développement des vraies dents ; de plus, structurellement différentes, elles avaient une morphologie et une fonction similaires.
Aujourd’hui, tous les oiseaux sont dépourvus de dents. Certains ont des denticules (appendices en forme de petite dent), mais ils n’affectent que la partie cornée du bec, la rhamphothèque. Depuis 66 millions d’années, et la disparition des dinosaures non-aviens et des oiseaux à dents du Crétacé, seuls des oiseaux sans dents ont peuplé notre planète. La perte des dents dans la lignée des oiseaux modernes a eu lieu il y a 110 ou 120 millions d’années, au cœur du Crétacé.
Des oiseaux marins géants uniques

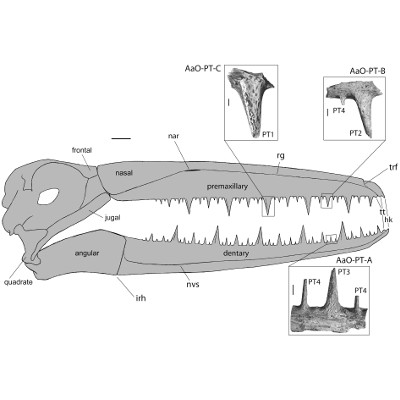
Les odontopterygiformes sont un groupe éteint d’oiseaux marins de grande taille : certains atteignaient 7 mètres d’envergure ce qui en fait les plus grands oiseaux volants connus (voir image 1). Ils ont vécu durant une grande partie du Cénozoïque, depuis il y a au moins 55 millions d’années, jusqu’à environ 2,5 millions d’années avant notre ère. Aussi, ils ne faisaient pas exception et possédaient un bec sans dents mais pourvu, cas unique chez les oiseaux, d’excroissances creuses en forme de dents aiguës et formées d’os appelées "pseudodents" (voir image 2). Ces formations étaient très certainement recouvertes de la rhamphothèque du bec du vivant de l’oiseau, celle-ci épousant ces formes coniques pointues.
De plus, ces pseudodents se répartissent sur les bords du bec de façon très particulière, formant des séquences alternant des pseudodents de grande taille, de petite taille et de taille intermédiaire de façon très régulière (voir image 3).
Un modèle pour expliquer la formation de dents osseuses

L’origine de ces pseudodents est longtemps restée très énigmatique, aucun équivalent actuel n’étant connu, même chez les autres vertébrés. Suite à une étude histologique antérieure de la même équipe (2013), des chercheurs de l’Institut de génomique fonctionnelle de Lyon (ENS de Lyon), en collaboration avec des chercheurs parisiens, proposent un modèle de développement de ces structures disparues, expliquant à la fois leur forme, leur composition et leur distribution spatiale. La précédente étude avait permis de s’orienter vers un développement relativement tardif des pseudodents, vers la fin de la croissance des os des mâchoires qui les portent. À ce stade, les pseudodents se développaient comme des excroissances osseuses le long des bords de chaque mâchoire. L’os de la mâchoire était donc au plus près de la couche externe du bec, l’épithélium, pas encore corné (kératinisé et durci). Cela suggère une interaction possible directement entre la couche externe de l’os et cet épithélium. Dans la formation d’une dent, l’épithélium oral interagit avec le mésenchyme (un tissus de soutient embryonnaire) et, lorsque la forme de la dent est obtenue, les tissus épithéliaux se minéralisent en émail et les tissus mésenchymateux en dentine. Dans le modèle proposé pour les pseudodents, l’épithélium est toujours capable d’envoyer des signaux développementaux, mais le mésenchyme n’est plus compétent : chez ces oiseaux, c’est la couche externe de l’os avant minéralisation qui "répond" et une forme conique de dent est obtenue, composée de matière osseuse.
Des champs morphogénétiques expliquent la distribution des pseudodents
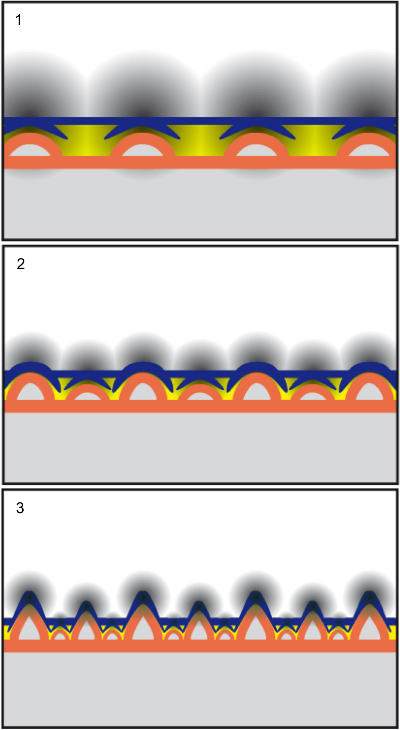
Restait à expliquer la distribution par taille des pseudodents le long des mâchoires. Des spécimens juvéniles trouvés ces dernières années au Venezuela et en Ukraine montrent que lors des premiers stades de croissance des pseudodents, celles qui seront les plus grandes sont les premières à croître et qu’elles finissent toutes leur croissance simultanément. Le plus probable est que des « champs morphogénétiques » contrôlant la croissance de chaque pseudodent empêchent celle des autres pseudodents à leur proximité. Toutefois, chaque champ d’inhibition a un rayon d’action qui décroît de jour en jour et, à un moment donné, le champ entre deux pseudodents devient libre pour permettre la croissance d’une autre à mi-chemin entre elles (voir image 4). L’analyse des spécimens connus de mâchoires portant des pseudodents, et les espacements entre celles-ci en fonction de leur taille, est compatible avec l’action de tels champs d’inhibition. Ce scénario explique la distribution unique de ces excroissances osseuses en fonction de leur taille. Il est intéressant d’observer que de vraies dents chez certains poissons (comme les requins-scies) présentent la même distribution de taille, suggérant un possible mode de contrôle de leur croissance homologue au modèle proposé chez les oiseaux géants.
Bricolage et co-adaptation
Il semble que la relative faiblesse de la mandibule des odontopterygiformes (absence de symphyse et présence d’un joint souple dans chaque branche) ait été compensée par la présence des pseudodents, aiguës, pour que l’efficacité soit garantie dans la capture et le maintien des proies (poissons, invertébrés marins…) dans la mâchoire. Les seules autres espèces connues partageant ces faiblesses étaient des oiseaux à dents du Crétacé, eux aussi piscivores.
Ainsi, la nature a trouvé une voie d’évolution qui apparaît comme un "bricolage", par l’utilisation détournée du programme de développement dentaire mais avec des cellules d’os en formation à la place des cellules du mésenchyme, aboutissant à la formation de structures différentes de vraies dents mais de fonction similaire. Grâce à une homologie profonde entre dents et pseudodents, ces dernières sont apparues en co-adaptation avec les caractéristiques mandibulaires des odontopterygiformes, permettant à ces oiseaux de garder une certaine efficacité dans la prédation. Cela leur a permis de vivre plus de 50 millions d’années sur presque tous les océans, leur extinction à la fin du Pliocène (2,5 millions d’années) restant une énigme.
Source : Louchart, A., Buffrénil, V. de, Bourdon, E., Dumont, M., Viriot, L. & Sire, J.Y. 2018. Bony pseudoteeth of extinct pelagic birds (Aves, Odontopterygiformes) formed through a response of bone cells to tooth-specific epithelial signals under unique conditions. Scientific Reports 8, 12952 (2018).
Glossaire :
Odontogénétique : L’odontogénétique est la discipline qui s’intéresse aux gènes impliqués dans l’odontogénèse, c’est-à-dire le processus de formation des dents.
Épithélium : Les épithéliums sont des tissus animaux (ensembles de cellules juxtaposées) qui servent notamment de tissus de revêtement.
Homologie : En Évolution, on parle d’homologie quand on retrouve chez deux espèces différentes des traits hérités d’un ancêtre commun.
Disciplines
Mots clés